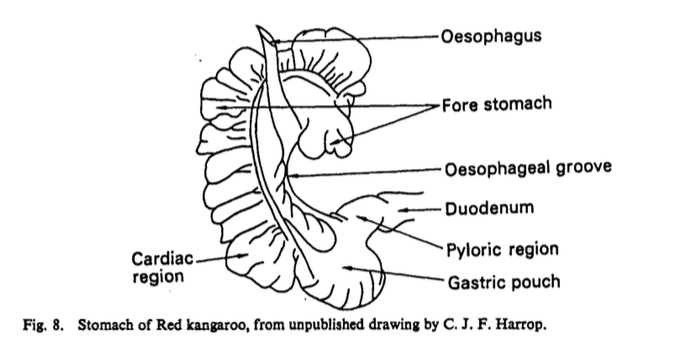
(Russell E.M., 1974).
|
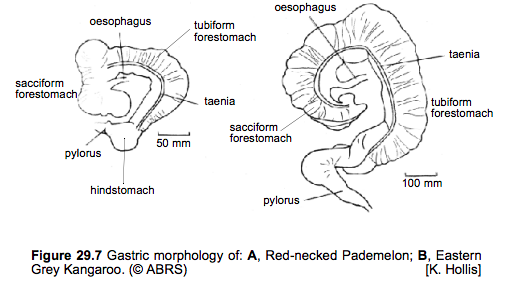
(Hume I.D. et al, 1989)
|
The parma wallaby (M. parma) is defined as a browser/grazer (Lentle R.G. et al., 2003) whereas the red kangaroo (M. rufus) is a grazer (Jarman P.J., 1984), however, both are classified as fore-gut fermenters which means a majority of fermentation of cell wall components occurs in the fore-stomach (Hume I.D., 1989). Foregut fermenters like the parma wallaby and the red kangaroo have developed larger and more differentiated fore-stomachs to accommodate for effective microbial fermentation to aid in the break down of hard cell wall components such as cellulose, hemicellulose and lignin (Hume I.D., 1989). Due to lack of information on the specific digestive anatomy of the parma wallaby, the red-necked pademelon (T. thetis) is used; because of its similar diet an habitat to the parma wallaby it can be assumed that these two species also have similar general digestive physiology.
The sacciform fore stomach, tubiform fore stomach regions which contain mucus secreting cells (Russell E.M., 1974) and the hind stomach region which contain acid and enzyme secreting cells (Russell E.M., 1974) make up the general structure of the macropodine stomach (Langer P., Dellow D.W., & Hume I.D., 1980). Microbial fermentation occurs mainly in the tubiform and sacciform (blind sac) regions of the fore stomach, with little occurring in the hind stomach, which is the main site of acid digestion (Hume I.D., 1989). Further microbial fermentation occurs in the colon in all foregut fermenters even when a ceacum is present (Hume I.D., 1989). The inner curvature of the fore stomach contains a "spiral groove analogous to the esophageal groove of ruminants," this allows non-fibrous food to by-pass fermentation in the fore stomach (Russell E.M., 1974). The fore stomach walls are separated into three taeniae which contain "semi-lunar fold between the taeniae" which form further haustrations; these structures allow for separation of solid and liquid digesta which is important in ensuring proper and efficient digestion of food (Langer P., Dellow D.W., & Hume I.D., 1980) These haustrations cause the stomach to have a colon-like appearance which is the most significant feature of the red kangaroos digestive tract (Hume I.D., 1989). In most macropodinae species the hind stomach makes up the smallest portion of the stomach and the sacciform and tubiform regions make up the majority of the stomach in varying amounts across species (i.e. in the T. thetis the sacciform is larger than the tubiform fore stomach whereas in the Eastern grey kangaroo the tubiform fore stomach was larger (Langer P., Dellow D.W., & Hume I.D., 1980)). The large tubiform region of the fore stomach in large kangaroos, such as the red kangaroo, results in lower retention times of digesta which means less time for fermentation to occur therefore poorer digestibility of plant cell wall components (Hume I.D., 1989). However, "marsupials chew food more thoroughly as they eat," so it might be possible that these animals rely more on chewing than fermentation to break down their food, but this is not clear (Hume I.D., 1989). Although the larger tubiform fore stomach in red kangaroos reduces their ability to ferment their food efficiently, it allows them to eat constantly for longer periods of time thus allowing them to rest in the shade when the days are too hot (Munn A.J., Dawson T.J. & McLeod S.R., 2010).
The sacciform fore stomach, tubiform fore stomach regions which contain mucus secreting cells (Russell E.M., 1974) and the hind stomach region which contain acid and enzyme secreting cells (Russell E.M., 1974) make up the general structure of the macropodine stomach (Langer P., Dellow D.W., & Hume I.D., 1980). Microbial fermentation occurs mainly in the tubiform and sacciform (blind sac) regions of the fore stomach, with little occurring in the hind stomach, which is the main site of acid digestion (Hume I.D., 1989). Further microbial fermentation occurs in the colon in all foregut fermenters even when a ceacum is present (Hume I.D., 1989). The inner curvature of the fore stomach contains a "spiral groove analogous to the esophageal groove of ruminants," this allows non-fibrous food to by-pass fermentation in the fore stomach (Russell E.M., 1974). The fore stomach walls are separated into three taeniae which contain "semi-lunar fold between the taeniae" which form further haustrations; these structures allow for separation of solid and liquid digesta which is important in ensuring proper and efficient digestion of food (Langer P., Dellow D.W., & Hume I.D., 1980) These haustrations cause the stomach to have a colon-like appearance which is the most significant feature of the red kangaroos digestive tract (Hume I.D., 1989). In most macropodinae species the hind stomach makes up the smallest portion of the stomach and the sacciform and tubiform regions make up the majority of the stomach in varying amounts across species (i.e. in the T. thetis the sacciform is larger than the tubiform fore stomach whereas in the Eastern grey kangaroo the tubiform fore stomach was larger (Langer P., Dellow D.W., & Hume I.D., 1980)). The large tubiform region of the fore stomach in large kangaroos, such as the red kangaroo, results in lower retention times of digesta which means less time for fermentation to occur therefore poorer digestibility of plant cell wall components (Hume I.D., 1989). However, "marsupials chew food more thoroughly as they eat," so it might be possible that these animals rely more on chewing than fermentation to break down their food, but this is not clear (Hume I.D., 1989). Although the larger tubiform fore stomach in red kangaroos reduces their ability to ferment their food efficiently, it allows them to eat constantly for longer periods of time thus allowing them to rest in the shade when the days are too hot (Munn A.J., Dawson T.J. & McLeod S.R., 2010).
